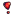The wonder molecule shares a structure similar to benzo(a)pyrene which is determined to be a mutagen. Benzo(a)pyrene is a molecule that when metabolised forms a mutagenic dihydrodiol epoxide which can form an adduct with DNA to alter the base pair coding and does this by binding to the N2 or the N7 of the guanine base. The molecule is able to do this as it is an intercalating agent, a flat planar molecule that can slip inbetween the DNA strands. Once it has bound to guanine it creates a space ((((VERIFY)))) which causes guanine to pair with adenine instead of cytosine. This change in base pair coding is carried forward to the next generation of cells where when the DNA strands are being read, it changes the G-C pair into an A-T pair which is linked to lung cancer due to the base-pair substitution.
Benzo(a)pyrene requires bioactivation via CYP1A1, 1B1. Some individuals may not express these enzymes which means they will not be affected by the molecule as they will not be able to metabolise it to the dihydrodiol epoxide metabolite which forms the adduct.
Due to having a similar structure to benzo(a)pyrene the wonder molecule should be subjected to several mutagenecity assays to determine if it is a mutagen like benzo(a)pyrene.
Short-term mutagenecity assays can determine if a molecule is a mutagen and also predict if it would require bioactivation or if it can act on its own. However, short-term mutagenecity assays cannot detect non-genotoxic carcinogens.
The Ames test is an example of an in-vitro mutagenecity assay that tests if a molecule is mutagenic. It is a bacterial reverse mutation assay that relies on different bacteria strains which have pre-existing mutations and any reversion to the wild type to determine if the added substance is mutagenic. The tester Salmonella typhimurium strain has an existing mutation in the histidine operon where it cannot create its own histidine and cannot form colonies in the absence of histidine. The test relies on the mutagenecity of the compound being tested to be able to mutate the mutated bacteria strain to revert them back to being able to produce histidine. The test is made robust enough to detect changes in base-pair or frameshift mutations.
The ames test has added layers to make it more sensitive in detecting mutagens such as an Rfa mutation which creates an LPS layer that coats the bacterial surface to increase the permeability of the cell wall increasing the penetration of bulky chemicals such as benzo(a)pyrene dihydrodiol epoxides and uvrB-bio deletion which reduces endogenous DNA repair mechanisms. The biotin gene is deleted to make the bacteria strain biotin dependent and cannot repair DNA on their own.
The test's results are easily seen with changes in colour of the plate wells. A change from purple indicator colour to yellow shows that the bacteria has mutated and began producing histidine which lowers the pH of the environment making it more acidic which is detected.
In an added layer to the ames test we can also test if the wonder molecule would require bioactivation like benzo(a)pyrene to be mutagenic. Humans have CYP450 enzymes which are able to metabolise polycyclic aromatic hydrocarbons into their active mutagenic forms and this dependency of metabolising enzymes can be tested by the addition of an S9 mix of a supernatant fraction of rat liver homogenate. Activity in S9 negative wells determine that a molecule would not require bioactivation and activity solely in S9 positive wells determine that a molecule would require bioactivation to produce mutagenic effects.
AN ACTION POTENTIAL TRAVELS ALONG THE MUSCLE FIBRE INTO THE SARCOLEMMA
THE ACTION POTENTIAL TRAVELS INSIDE THE SARCOLEMMA INTO THE T TUBULES
THE T TUBULES ARE SITUATED NEXT TO THE SARCOPLASMIC RETICULUM VIA A SERIES OF PROTEINS
INSIDE THE SARCOPLASMIC RETICULUM, THERE ARE TONS OF CA2+ IONS
THE ADJACENT SARCOPLASMIC RETICULUM CONNECTED TO THE T TUBULES IS CALLED A TERMINAL CISTERNAE AND THEY MAKE UP A TRIAD WITH THE T TUBULES
WHEN AN ACTION POTENTIAL TRAVELS ALONG A T TUBULE, A VOLTAGE SENSITIVE GATE OPENS AND CHANGES SHAPE CAUSING THE CALCIUM CHANNEL TO OPEN UP AND RELEASE CALCIUM TO THE SARCOPLASM
THE CROSS BRIDGE CYCLE
WITH CALCIUM IN THE SARCOPLASM, THEY CAUSE TROPONIN TO CHANGE SHAPE TO EXPOSE THE ACTIN MYOFILAMENTS
THE MYOSIN HEADS CAN BIND TO THE ACTIN MYOFILAMENTS ONLY IF IT IS ACTIVATED
IT IS ACTIVATED WHEN ATP BINDS TO THE HEAD AND IS HYDROLYSED TO ADP AND INORGANIC PHOSPHATE WHICH ARE BOTH RELEASED. THE HYDROLYSIS CAUSES THE HEAD TO ENTER INTO A COCKED POSITION AND READY TO BIND TO THE ACTIN HEAD.
ONCE ADP IS RELEASED, THE MYOSIN PIVOTS TOWARDS THE CENTRE AND THE MYOSIN HEAD ATTACHES.
THE DETACHMENT OCCURS WHEN ANOTHER ATP MOLECULE BINDS TO THE MYOSIN HEAD AND IT IS RELEASED FROM THE CROSS BRIDGE
LETS SEE BRAIN SO SYNAPSES
ACTION POTENTIALS TRAVEL ALONG THE AXON TERMINAL AND PROPAGATES THE VOLTAGE GATED CALCIUM CHANNELS TO OPEN
CALCIUM IONS ENTER INTO THE TERMINAL AND GO TO THE SYNAPTIC VESICLES WHICH RELEASE ACETYLCHOLINE
ACETYLCHOLINE BINDS TO A RECEPTOR IN THE SYNAPTIC TERMINAL AND ALLOWS THE LIGAND GATED CHANNELS TO OPEN
NA+ INFLUX, K+ EFFLUX CAUSING THE MEMBRANE POTENTIAL TO BE LESS NEGATIVE
ONCE THRESHOLD IS REACHED, ACETYLCHOLINE IS RELEASED VIA EITHER EXOCYTOSIS OR DIFFUSION AND tada
SALT CHANNELS UM UM I KNOW ABOUT THIS
NaK ATPase (or sodium pump) allows for 3Na+ to exit the basolateral membrane while allowing 2K+ into the membrane
NAK2Cl symporter allows in 2Cl-, Na+, K+ into the basolateral membrane
Cl- exits out via passive diffusion through an ion channel to the apical membrane surface to the lumen of the gut
K+ exits out via its channel
and the Na+ that enters exits via the sodium pump (which creates the force required in the first place for secondary active transport)
Cl- leaving induces paracellular Na+ and H2O fluxes